

Учебная работа. Статья: О некоторых стратегических ошибках в современном креационном мышлении
Хоменков А. С.
широко известна история, как известный популяризатор эволюционной доктрины Эрнст Геккель подделал в свое время рисунки эмбрионов. Геккель пытался увеличить сходство между эмбрионами человека и эмбрионами различных видов живых существ и, тем самым, подчеркнуть родственную эволюционную связь человеческого племени с представителями разнообразных рыбьих, птичьих и собачьих племен.
Исторический путь идей Геккеля
В дальнейшем стиль мышления Геккеля стал основой для пропаганды эволюционных идей в школьных и университетских учебниках биологии. Все когда-то знакомились в школе с рисунком человеческого эмбриона, помещенном в учебнике «Общая биология» по соседству с рисунком эмбриона рыбы. И там и тут были тщательно вырисованы так называемые «жаберные щели», а в тексте давались пояснения по поводу того, что человеческий эмбрион имеет ряд признаков, общих с эмбрионом рыбы, и что это является доказательством того, что эволюционные предки человека были с жабрами. Отсюда делался вывод, что в эмбриональном развитии человека можно найти следы его эволюционной истории. Чарльз Дарвин, кстати, объявил в свое время это «открытие» Геккеля главным доказательством своего эволюционного учения (Головин, 2000, с. 36). «Теория эмбриональной рекапитуляции» Геккеля стала в последующие годы с фантастической скоростью распространяться по всему миру, стимулируя при этом распространение абортов. Ведь если человеческий зародыш – еще не человек в собственном смысле этого слова, а некое неведомое существо с чем-то вроде рыбьих жабр и собачьего хвоста, который Геккель также смог «увидеть» на своих препаратах, то его не жалко и убить. Рисунки эмбрионов в школьных учебниках были призваны подтвердить эту точку зрения, и в этих рисунках сходство с «рыбьими признаками» человеческих эмбрионов особо подчеркивалось. Сторонники идеи Творения заняли противоположную позицию и старались при всяком удобном случае указать на фальсификации Геккеля и его последователей и подчеркивали различия в течение эмбриогенеза человека и рыбы. Перед неискушенным же читателям возникал определенный стереотип понимания сути дискуссии: чем больше можно было бы найти «рыбьих признаков» в человеческом эмбрионе, тем больше шансов на подтверждение своей правоты у сторонников эволюционной точки зрения. И наоборот: чем меньше существует там этих рыбьих признаков, тем большая вероятность того, что правы креационисты.
однако эта проблема не так проста. более того, каким бы странным это не могло кому-то показаться, она, скорее, решается с точностью до наоборот. Сходство в эмбрионах рыбы и человека указывает именно на правоту креационистов. впрочем, не будем забегать вперед и для начала расскажем о том, что говорит современная наука об «открытом» Геккелем «законе эмбриональной рекапитуляции».
«Синтез германского романтизма и дарвинизма»
Если мы откроем современный фундаментальный трехтомный труд «Биология развития», написанный ученым с мировым именем Скоттом Ф. Гилбертом, то увидим, что «биогенетический закон» характеризуется здесь самым нелестным образом. Автор называет его «синтезом германского романтизма и дарвинизма» (1993 (Т. 1), с. 146) и сфабрикованным «гибельным союзом эмбриологии и эволюционной биологии» (1995 (Т. 3), с. 309). термин «германский романтизм» здесь появился из-за того, что Геккель, как известно, жил в Германии. Ну а указание на то, что союз эмбриологии и эволюционной биологии был сфабрикован, является напоминанием о мошенничестве Геккеля. В целом же, анализируя это «открытие» Геккеля, Гилберт пишет, что его точка зрения на процесс эмбрионального развития «была дискредитирована даже раньше, чем… предложена» (1993 (Т. 1), с. 146). здесь имеется в виду критика знаменитым эмбриологом Карлом Бэром доэволюционных форм того стиля мышления, который впоследствии «оседлал» Геккель. Бэр фантазировал, что могло бы получиться, если бы птицы стали писать учебники эмбриологии. Вот что мы смогли бы тогда прочесть в этих учебниках: Эти четверо- и двуногие животные во многом сходны с нашими собственными зародышами: кости их черепа не сращены; у них нет клюва, так же, как и у нас в первые пять или шесть дней насиживания; все их конечности очень сходны друг с другом, как и у нас в тот же самый период; на их теле нет ни одного настоящего пера, а лишь очень тонкие стержни, так что когда мы, уже оперившись, сидим в гнезде, мы уже достигли в своем развитии такого уровня, какого им никогда не удастся достичь… И эти-то млекопитающие, которые в течение столь долгого времени после появления на свет не в состоянии сами добывать себе пищу, которые никогда не смогут оторваться от земли, считают себя более высокоорганизованными, чем мы?» (цит. по: Гилберт (Т. 3), 1995, с. 310).
однако самое главное в этой проблеме заключается в том, что, как указывает Гилберт, в основе «биогенетического закона» «лежат ложные предпосылки» и эти ложные предпосылки связаны не с додарвиновскими работами Карла Бэра, но выявились в результате достижений современной науки (1993 (Т. 1), с. 146).
Гилберт, кстати, стоит на эволюционных позициях и его свидетельство в этом плане обладает особой ценностью. Подобные же свидетельства можно найти и у других ученых, активно занимающихся защитой эволюционной гипотезы. Так, знаменитый ученый-эволюционист Джордж Симпсон писал еще в 1965 году следующее:«В наше время достоверно установлено, что онтогенез не повторяет филогенез» (цит. по: Хэм и др., 2000, с. 115).
Этот вывод вытекает из конкретных положений современной биологии, связанных с целым ее отраслей. В частности, об этом свидетельствует гистология. Так, основоположник школы российской гистологии академик А.А. Заварзин пришел в свое время к отрицанию возможности приложения «биогенетического закона» к гистологической организации живых организмов. Этот вывод он сформулировал после долгих поисков примеров «рекапитуляций» и «высказал столь резко свою точку зрения только в конце своего творческого пути. “Не только я, но и все, кто интересовался приложением эволюционной теории к тканям, – свидетельствует Заварзин, – не могли, конечно, не искать примеров рекапитуляций в развитии тканей”. И признается, что “это оказалось далеко не легкой задачей”. огромный материал по эволюционной динамике нервной системы, крови и соединительной ткани не содержит, по Заварзину, ни одного примера рекапитуляции. Примеры гисторекапитуляций, приводимые другими авторами, он считал неубедительными» (Мирзоян, 1980, с. 93). Академик Заварзин, кстати, указывал в более широком плане, что «гистологические факты не укладываются в филогенетические схемы, созданные сравнительной анатомией» (Мирзоян, 1980, с. 65).
По словам другого известного российского гистолога – Н.Г. Хлопина – «биогенетический закон, которым широко пользовался Геккель при построении своей филогенетической системы, не оправдал себя в области гистологии» (цит. по: Мирзоян, 1980, с. 150). Согласно взглядам этого ученого, «трактовка Геккелем ранних этапов эмбрионального развития под углом зрения эволюции тканей несостоятельна» (цит. по: Мирзоян, 1980, с. 150).
Итак, два выдающихся ученых-гистолога указывают на несостоятельность «биогенетического закона». Два других ученых-эволюциониста с мировым именем – Р. Рэфф и Т. Кофмен – поясняют, что несостоятельность «закона эмбриональной рекапитуляции» вытекает из трех фундаментальных положений современной биологии – менделевской генетики, обособленности клеток зародышевой линии и важности морфологических признаков на протяжении всего зародышевого развития. По словам этих ученых, эти три фундаментальные положения современной биологии «положили конец рекапитуляции» (Рэфф, Кофмен, 1986, с. 31). И этот конец «биогенетическому закону» положен довольно давно. По словам профессора Кейта Томпсона из Йельского университета, «его изгнали из учебников биологии в 50-х годах, а как тема серьезного теоретического исследования он перестал существовать еще в 20-е годы» (цит. по: Хэм и др., 2000, с. 115). Имеется в виду ХХ столетие. В настоящее же время, как пишет профессор Кейт, «биогенетический законнесмотря на титанический труд нескольких поколений ученых-эволюционистов и пестрый спектр мнений по вопросу интерпретаций эмбриологических фактов, общепризнанное эволюционное объяснение эмбриологических закономерностей отсутствует. Заведующий кафедрой теории эволюции биологического факультета Московского Государственного университета профессор А.С. Северцов пишет, что «можно говорить лишь о закономерностях эволюционной перестройки онтогенеза» и что эта перестройка есть «одна из важнейших, сложнейших и интереснейших проблем биологии» (Северцов, 1987, с. 222).
таким образом, современная биология наложила запрет на геккелевский стиль мышления – и это признают даже ученые-эволюционисты. То, что этот стиль продолжает существовать в современных учебниках биологии и различных популярных брошюрах – это проблема уже не биологическая, а, скорее, социально-психологическая и ее разбором мы сейчас заниматься не будем. Нас в данный момент интересует другой вопрос: существует ли сходство эмбриона человека и эмбриона рыбы, и если оно существует, то каков его смысл?
Сходство все же существует
В современных учебниках по эмбриологии вместо термина «жаберные щели» иногда употребляются термины «фарингеальные дуги» или «трещины». Тем самым подчеркивается, что эти образования никогда не выполняют у человека функций, связанных с дыханием, и превращаются в процессе эмбрионального развития в совершенно иные структуры, не имеющие отношение к дыхательной функции. Но, в то же время, даже креационисты рассматривают эти образования как «внешне напоминающие “жаберные щели” у эмбриона рыб» (Хэм и др., 2000, с. 116). другой случай внешнего сходства эмбриона человека и эмбриона рыб – выраженное метамерное деление тела или сегментация. В какой-то степени такое сегментированное, метамерное деление можно обнаружить и у взрослого человека, особенно в области его грудной клетки – ребра расположены параллельными рядами и соединены с позвоночником, состоящим из отдельных позвонков. Но это все же слабый отголосок того, что наблюдается у человеческого эмбриона. Там сегменты тела ярко выражены и напоминает то, что мы можем наблюдать у рыб не только у эмбриональной, но и у взрослой их стадии. У креационистов существует попытка объяснить это явление с функциональных позиций: сегментация имеет конструктивное такой подход неоправданной данью непомерно разросшейся в свое время тенденции объяснять все морфологические закономерности с позиции исторического и функционального подхода? Такой подход является методологически не оправданным (Канаев, 1966, с. 199), а его использование креационистами особенно абсурдно по той причине, что он органически гораздо более связан с эволюционным стилем мышления и редукционизмом (Светлов, 1982, с. 50), чем типологический подход «идеалистической морфологии». Попробуем подойти к проблеме существования метамерного деления тела эмбриона человека и наличия в нем «фаренгиальных дуг» на основании того варианта типологического подхода, который разрабатывался автором в статье «Возможна ли теоретическая биология?» (Хоменков, 2005).
«Матрицы жизни» и проблемы эмбриогенеза
Напомним, что для адекватного описания ряда биологических закономерностей, мною были предложены формальные схемы – «матрицы жизни», являющиеся неким аналогом того, чем является математический аппарат при решении проблем физических.
Матрица представляет собой довольно простую схему, состоящую из ряда расположенных друг над другом горизонтальных линий, каждая из которых является неким интегральным показателем свойств какого-либо вида живых существ, связанных с его таксономическим положением. Нулевой уровень характеризует свойства общебиологического характера, первый – свойства, характеризующие то царство, к которому принадлежит рассматриваемый нами вид, второй уровень – свойства типа, третий – подтипа и т.д., вплоть до последнего уровня, в котором отражены видовые свойства (рис.1).
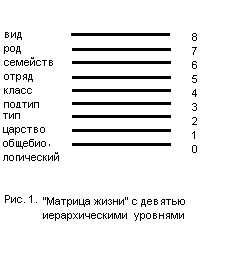
Манипулируя содержанием формальных схем, можно создавать модели того, что мы наблюдаем в природе в случае таких явлений как «параллелизм», «конвергенция», мозаичные формы и ряда других биологических явлений, традиционно объясняемых с эволюционных позиций (Хоменков, 2005). Эффективность таких схем подразумевает наличие разумного замысла, лежащего в принципах мироустроения. Аналогичным образом дело обстоит и в сфере физической реальности, которая, по свидетельству исследователей, подчинена принципу математической простоты и симметрии.
Но может ли в таком случае использование формальных схем – «матриц жизни» – помочь нам в объяснении тех закономерностей эмбрионального развития организмов, которые были описаны еще Карлом Бэром и которые Эрнст Геккель использовал в качестве «доказательства» эволюции? Может ли этот метод объяснить, почему человеческий зародыш имеет общие черты с зародышем рыбы, черты, которые гораздо ближе к рыбьим, чем к человеческим?
Очевидно, что если наши формальные схемы являются отражением разумного замысла Создателя живой природы, то процесс индивидуального развития организма должен иллюстрироваться этими схемами, как «заполнение» их уровней начиная от нулевого – общебиологического, и кончая восьмым – видоспецифичным, то есть «снизу вверх».
здесь уместно проведение некоторой аналогии со сферой технической деятель человека. Если мы, к примеру, хотим сконструировать серию автомобилей различного типа, то этот процесс логично начать с самых общих принципов, которые свойственны абсолютно всем автомобилям, например, положить в основу нашей конструкции платформу с четырьмя (как минимум) колесами и систему управления с находящимся спереди рулем. Далее на основании этих общих принципов следует подумать о размерах автомобилей, мощности мотора, форме кузова и т.д. – то есть о тех принципах, которые характерны для определенного класса машин. И лишь в самом конце можно позаботиться о второстепенных деталях, связанных с комфортностью использования каждого конкретного вида автомобилей.
Подобное конструирование будет основываться на иерархической шкале, которая очень напоминает наши «матрицы». Очевидно, что с чем-то похожим мы должны столкнуться и в процессе эмбриогенеза, если, конечно, подходить к этому процессу со стороны представлений о разумном замысле, лежащем в основе мироустроения. Процесс индивидуального развития живых организмов должен проходить путем появления сначала общих признаков, и лишь в конце – самых специфических, то есть «снизу вверх». Совпадает ли это теоретическое ожидание с тем, что мы наблюдаем в реальной жизни?
Уже упоминаемый нами выдающийся эмбриолог Карл-Эрнст фон Бэр (1792 — 1876), писал: «Я зафиксировал в спирте двух маленьких зародышей, забыв пометить их. А теперь не могу определить, к какому роду животных они относятся. Они могут быть ящерицами, мелкими птицами или даже млекопитающими!» (цит. по: Гилберт, 1993 (Т.1), с. 145).
Эта неразличимость очень хорошо видна на примере наших формальных схем. заполнение уровней «матриц» с нулевого по третий (что соответствует ранним этапам эмбриогенеза), у представителей перечисленных Бэром классов позвоночных должно представлять весьма сходный процесс. Ведь согласно Бэру «история развития особи есть история растущей во всех отношений индивидуальности» (Лункевич, 1960, с. 382). Бэр писал: «зародыш определенной животной формы вовсе не пробегает через ряд других определенных же форм, а скорее отделяется от них» (цит. по: Лункевич, 1960, с. 382), т.е. отходит все дальше и дальше от всех форм, кроме той, к которой направлено его собственное развитие. Что же касается более детальной трактовки открытых им особенностей индивидуального развития, то Бэр, подытоживая результаты своих наблюдений, выделил в свое время следующие основные законы: «1) признаки, общие для большой группы животных, образуются у зародыша раньше, чем частные признаки; 2) из более общего в соотношении форм образуется все более частное и затем наиболее специализированные признаки; 3) каждый зародыш определенной животной формы не проходит стадии развития других животных форм, а все более отделяется от них;4) зародыш высшей формы никогда не бывает похож на другую форму, а лишь на ее зародыш» (цит. по: Северцов, 1987, с. 194–195).
характерно, что Бэр полагал, что «обнаружил божественный план, по которому развиваются все организмы в такой группе, как позвоночные» (Гилберт, 1993 (Т. 1), с. 147). И при всем этом его законы до сих пор не потеряли своей актуальности, хотя из них было изъято то метафизическое содержание, которое в них вкладывал этот великий эмбриолог. Как подчеркивает профессор А.С. Северцов, в схеме Карла Бэра «два основных положения, получившие название принципа зародышевого сходства и принципа специализации индивидуального развития, сохранили свое значение до настоящего времени» (Северцов, 1987, с. 195). Сам Бэр писал по поводу этих принципов следующее: «Чем более несходны друг с другом две животные формы, тем дольше в глубь истории развития (эмбрионального) нужно погрузиться для того, чтобы найти между ними сходство» (цит. по: Лункевич, 1960, с. 381).
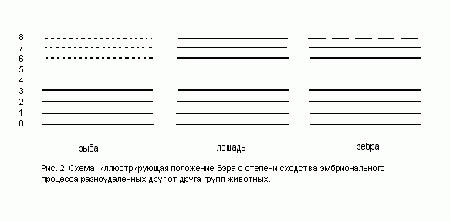
характерно, что эта закономерность настолько явно вытекает из свойств нашей формальной схемы «матриц», что эти формальные схемы можно считать ее прямыми иллюстрациями. Этот факт можно проиллюстрировать следующим рисунком, на котором показаны три «матрицы жизни» – лошади, зебры и рыбы (рис. 2). Чтобы найти общее в эмбрионах первых двух живых существ, следует «погрузиться» до второго (сверху) уровня «матрицы», который соответствует уровню рода. чтобы найти общее в «матрицах» лошади и рыбы – нужно дойти до шестого сверху уровня (подтипа), то есть до гораздо более ранних стадий развития (вспомним, процесс эмбриогенеза – это последовательное «заполнение» матриц, идущее «снизу вверх»).
однако каким образом с помощью наших формальных схем можно объяснить те многочисленные факты, которые традиционно привлекаются в качестве иллюстрации «биогенетического закона»? Почему зародыш человека имеет определенную степень схожести с зародышем рыбы? чтобы ответит на эти вопросы, нужно дополнить нашу формальную схему «матриц» некоторыми конкретными гистологическими данными.
Типологический ключ к «главному доказательству эволюции»
Некоторые ученые, размышлявшие о том, каким образом можно объективно оценить степень сложности какого-либо организма, пришли к выводу, что наилучшим показателем здесь может служить «число типов клеток, различаемых у представителей данной группы» (Рэфф, Кофмен, 1986, с. 327) . У представителей разных систематических групп этот показатель будет весьма различен. «Так, у бактерий имеются клетки двух типов (вегетативные клетки и споры), у дрожжей – 3–4 типов, у водорослей и грибов – примерно 5 типов, у губок – 11, у кишечнополостных – 14–20, у растений – от 20 до 40, у кольчецов – примерно 55» (Рэфф, Кофмен, 1986, с. 327). Для более высокоорганизованных животных такие цифры получить сложнее, но, тем не менее, можно давать хотя бы приблизительные цифры. У разных групп рыб количество клеток по приблизительным оценкам колеблется в пределах 70–80. Что же касается млекопитающих, то у них количество типов клеток как минимум 100, но, по мнению некоторых ученых, на целый порядок больше (Рэфф, Кофмен, 1986, с. 327–328). Приведенные цифры можно считать объективным выражением той возрастающей сложности «лестницы живых существ» о которой писал в свое время еще Аристотель . И эта возрастающая сложность имеет свое выражение в конкретных гистологических данных: «чем проще общая организация животных, тем более универсальны и мультифункциональны ткани. И наоборот, чем выше и сложнее морфологическая организация, тем специфичнее организация гистоструктур и тем больше их разнообразие» (Мирзоян, 1980, с. 132).
Каким же образом эту закономерность и приведенные выше цифры можно отобразить на наших «матрицах»? чтобы дать конкретный ответ на этот вопрос, изобразим «матрицы» двух классов позвоночных животных – костных рыб и млекопитающих.
Очевидно, что нижние четыре уровня «матриц» (от общебиологического уровня до уровня подтипа – Vertebrata) как рыб, так и млекопитающих, должны быть идентичными. Разница, существующая в гистологической структуре этих двух таксономических групп, должна формироваться за счет пяти верхних уровней «матриц». «Как эмбрионы переходят от общих признаков к более частным, так и в процессе тканевой дифференцировки сначала появляются признаки общих тканевых типов, затем – признаки специализированных групп тканей и, наконец, признаки отдельных гистологических структур» (Мирзоян, 1980, с. 220–221). И, при этом, «разнообразие и специфичность тканевых производных каждого зародышевого листка возрастает по мере повышения организации животных» (Мирзоян, 1980, с. 226). На схеме это явление будет запечатляться следующим образом (рис. 3). У «матрицы» рыб верхние пять ступеней должны быть гораздо эже аналогичных ступеней млекопитающих, что должно уменьшить общую длину всех ступеней представителей класса рыб.
теперь посмотрим, какое различие в прохождении эмбрионального процесса рыб и млекопитающих будет следовать из особенностей скорректированных за счет гистологических данных «матриц». Когда процесс эмбрионального развития пройдет немного менее половины своего пути (что соответствует заполнению четырех нижних уровней «матриц»), зародыши рыб и млекопитающих должны в весьма значительной степени походить друг на друга и более походить на взрослую рыбу, чем на взрослое млекопитающее. последнее вытекает из того, что заполненные четыре нижних ступени «матриц» рыбы содержат в себе уже больше половины признаков взрослой особи этого класса, в то время как у млекопитающих этот этап эмбриогенеза – меньше половины. другими словами – на ранних стадиях эмбриогенеза определенные «рыбьи черты» должны быть в какой-то степени свойственны всем представителям позвоночных потому, что рыба является наиболее просто организованным представителем этого подтипа и ее «конструктивная идея», как бы, лежит в основе строения всей этой таксономической группы.
здесь поневоле вспоминаются слова известного французского ученого XVIII века Жоржа Бюффона, писавшего, что «Высшее Существо создавая животных, пожелало употребить лишь одну идею варьируя ее в то же время самым различным образом, дабы человек мог равным образом восхищаться великолепием исполнения и простотой замысла» (цит. по: Канаев, 1963, с. 35).
И все же человек всегда остается человеком
Итак, определенные общие признаки у эмбрионов рыб и млекопитающих могут являться отражением той тонкой типологической организации живой материи, которая лишь весьма грубо и схематично может быть отражена на схеме наших «матриц».
Характерно, что Бэру и его современникам было известно о «жаберных щелях» куриных эмбрионов. Однако этот факт, как пишут современные исследователи «трактовался Бэром в духе идеалистической морфологии с ее поисками “плана строения” и архетипа, в духе представлений Кювье о едином плане строения организмов одного типа, отличном от планов строения других типов. Согласно представлениям додарвиновской эмбриологии, индивидуальное развитие идет от общего к частному, т. е. наличие жаберных щелей есть общий признак эмбрионов всех позвоночных, а не свидетельство прохождения предками птиц рыбообразной стадии» (Воронцов, 1999, с. 315–316). Сам Бэр, отрицая возможность уподобления зародышей каким-либо взрослым формам, писал, что «зародыш у высших животных на каждой стадии своего развития должен обнаруживать сходство не с той или иной деталью строения взрослых форм, а с их общей сущностью» (цит. по: Воронцов, 1999, с. 248).
Эту «общую сущность» для разных стадий развития разных классов позвоночных животных, если обратиться к схеме наших «матриц», будут олицетворять нижние ступени схемы, с «прохождения» которых начинается эмбриональное развитие любого представителя этого подтипа. Что же касается некоторых деталей строения, то здесь мы можем обнаружить и существенные различия между зародышами разных групп животных. В частности, исследователи отмечают различие в деталях строения человека и макаки-резуса уже на стадии оплодотворенной яйцеклетки (Юнкер, Шерер, 1997, с. 129). «Это означает: основы строения тела позвоночных животных закладываются абсолютно разными способами» (Юнкер, Шерер, 1997, с. 129). Проявляющееся же в дальнейшем сходство являются иллюстрацией того единого гиперплана, в соответствии с которым было «спроектировано» все многообразие биосферы, таким образом, что минимальным количеством «мазков» Творец «изобразил» на «холсте мира» максимально возможное разнообразие живых существ. Но использование «общих мазков» в творческом процессе не исключает существование глубокой индивидуальности в каждом сотворенном роде, индивидуальности, проявляемой с первых моментов эмбриогенеза в виде характерных деталей развития различных эмбрионов. Последнее положение не отражается на наших формальных схемах. Мы должны помнить, что эти «матрицы жизни», как и любые другие модели, не могут абсолютно адекватно отразить реальность, и в них отражены лишь самые общие принципы организации живой природы. Их использование наверняка даже более ограничено, чем использование математического формализма при описании физических законов. опираясь на эти формальные схемы, мы вряд ли сможем сделать открытие, аналогичное открытию планеты Плутон «на кончике пера». Но, в то же время, эти формальные схемы могут быть определенной иллюстрацией, поясняющей уже известные биологические факты и позволяющие глубже проникнуть в сущность биологических явлений.
Итак, наличие «общего знаменателя» в явлении эмбриогенеза разных систематических групп не исключает существования и глубоких индивидуальных различий. Здесь следует напомнить традиционное христианское представление, поднятое в предыдущей публикации на это тему (Хоменков, 2005, с. 289–292). Бытие каждого явление нашего мира определяется его запредельным, идеальным «корнем» – логосом, являющимся энергией Ипостасного Логоса. Этот логос, направляет индивидуальное развитие каждого существа по своему особому пути, несмотря на то, что наблюдаются и определенные общие детали в развитии разных биологических объектов, связанные с тем, что логосы разных живых существ, являясь некими «разветвлениями» Логоса Ипостасного, должны содержать в себе некоторую часть общей информации. Но эта схожесть не перечеркивает связанной с запредельными «корнями» каждого живого существа индивидуальности. И человек в этом строго направленном его логосом развитии всегда останется человеком, а рыба всегда останется рыбой.
Откуда берутся неоправданные стереотипы?
Итак, существует запрет со стороны теоретических положений современной биологии на «закон эмбриональной рекапитуляции», связанный с именем Геккеля. В то же время существуют эмпирически наблюдаемый факт схожести эмбрионов различных классов позвоночных и факт определенной близости этой общей эмбриональной стадии к морфологии самого «простого» класса позвоночных – рыбам. И этот факт очень хорошо трактуется с позиции типологического подхода, свидетельствующего о разумном замысле, положенном в основу многообразия жизненных форм, населяющих землю.
Мог ли подозревать Эрнст Геккель, когда он занимался подтасовкой эмбриологических данных, что те свидетельства, значимость которых он таким образом пытался преувеличить, в конце концов перестанут соответствовать эволюционному учению, но станут однозначным подтверждением представлений о Творце мира, положившего в основу всего творения единый гиперплан?
Впрочем, не менее интересен и другой вопрос: почему в современном креационизме вместо творческого осмысления проблем эмбриогенеза присутствует некий «негативный снимок» Геккелевского стиля мышления? То, что для Геккеля и его последователей является белым, для современных креационистов стало черным, но общие очертания картины при этом меняются очень мало.
Очевидно, что в современном креационном движении недостаточно присутствует позитивный элемент, связанный с представлениями о Божественном замысле, положенным в основу творения, и проявляющимся в данном случае в закономерностях эмбриогенеза. Точно такое же игнорирование замысла Творца наблюдается, кстати, и в вопросе, связанном с полемикой между эволюционистами и креационистами по поводу генетической и биохимической дистанции между человеком и человекообразными обезьянами. В этой области сложилась точно такая же ситуация, как и в эмбриологии. Ученые-эволюционисты стремятся подчеркнуть генетическую и биохимическую близость между человеком и обезьянами, связывая этот факт с эволюционной близостью между ними. Ученые-креационисты, наоборот, при всяком удобном случае приводят те данные, согласно которым дистанция между этими видами не такая уж близкая (смотрите: Хэм и др., 2000, с. 110–116). В то же время, если вспомнить традиционное христианское представление, что обезьяна создана как пародийный двойник человека, то смысл уникальной генетической и биохимической близости становится понятным: за обезьяньей внешностью скрывается «начинка» настолько близкая к человеку, что мы своим «шестым чувством» угадываем себя в этом существе (подробнее: Хоменков, 2001а, 2001б).
характерно, что когда было выяснено наличие феноменальной биохимической и генетической схожести между человекообразными обезьянами и человеком (схожесть на уровне подвидов!), некоторые ученые выдвинули предположение, что подобная же схожесть наблюдается и во внешнем виде, но мы ее не замечаем, так как человек субъективно более склонен улавливать различия между собой и обезьяной. Было высказано предположение, что «таксономисту, не относящемуся к Homo sapiens, шимпанзе и человек казались бы видами-двойниками, принадлежащими к одному роду» (Рэфф, Кофмен, 1986, с. 92–93). Это предположение было подвергнуто объективной проверке: проведены «количественные сравнения между человеком и шимпанзе по таким же морфологическим признакам, которые используются систематиками для выявления различий между бесхвостыми амфибиями по форме их тела. Оказалось, что человек и шимпанзе различаются по этим критериям в несколько большей степени, чем бесхвостые амфибии, принадлежащие к разным подотрядам» (Рэфф, Кофмен, 1986, с. 93). Перед нами случай, кажется, более не повторяемый в живой природе: сходство по внешности на уровне подотряда, а сходство по «внутренней начинке» на уровне подвида! Это явное указание на разумный замысел, на те уникальные штрихи сотворившего наш мир Художника, которыми Он подчеркнул особое положение в этом мире человека и его пародийного двойника.
Еще в VII веке христианский подвижник и философ св. Максим исповедник писал, что Бог, по созданному им тварному миру «познается, как художник по произведению», при этом, познается «путем научного изучения и познания творений» (Максим Исповедник, 1994, с. 245). В современном креационном мышлении ощущается явная недооценка силы этого мировоззренческого положения, недооценка, приведшая к неоправданным стереотипам в понимании реальности сотворенного мира.
Список литературы
Вильвовская А.В. человек и космос. – М.: Наука. 1994.
Воронцов Н.Н. Развитие эволюционных идей в биологии. – М.: Издательский отдел УНЦ ДО МГУ, прогресс — Традиция. 1999.
Гилберт С. Биология развития. В 3-х томах. – М.: мир. 1993–1995. Т. 1–3.
Головин С. Эволюция мифа. – М.: Паломник. 2000.
Догель В.А. Сравнительная анатомия беспозвоночных. – Л.: 1938. Ч. 1.
Канаев И.И. Очерки из истории сравнительной анатомии до Дарвина. – М. – Л.: Изд. АН СССР. 1963.
Канаев И.И. Очерки из истории проблемы морфологического типа от Дарвина до наших дней. – М. – Л.: Наука. 1966.
Лункевич В.В. От Гераклита до Дарвина. – М.: Изд. мин. просвещения РСФСР. 1960. Т. 2.
Максим Исповедник. Комментарии // В кн.: Дионисий Ареопагит. О Божественных именах. О мистическом богословии. – СПб.: Глагол. 1994.
Мигдал А.Б. Физика и Философия // Вопросы философии. – 1990. №1.
Мирзоян Э.Н. Развитие основных концепций эволюционной гистологии. – М.: Наука. 1980.
Рэфф Р.; Кофмен Т. эмбрионы, гены и эволюция. – М.: Мир. 1986.
Северцов А.С. Основы теории эволюции. – Изд. МГУ. 1987.
Светлов П.Г. (под редакцией) Александр Александрович Любищев. Л.: Наука. 1982.
Хоменков А.С. Почему некоторые обезьяны человекоообразны? // Божественное Откровение и современная наука. Вып. 1. М.: Паломник. 2001 // православная беседа. 2001. №№ 3–4.
Хоменков А.С. Возможна ли теоретическая биология? // Православное осмысление творения мира. Сборник докладов конференции XIII Международные Рождественские Образовательные чтения. М.: 2005.
Хэм К., Сарфати Дж., Виланд К. книга ответов. – Сиферополь: Изд. Христианского научно-апологетического центра. 2000.
Юнкер Р.; Шерер З. История происхождения и развития жизни. – СПб.: Кайрос. 1997.